호르몬 & 행동(Hormones & Behavior)
호르몬 & 행동
By Randy J. NelsonThe Ohio State University
이 모듈의 목표는 호르몬과 행동에 대한 주제를 소개하는 것입니다. 이 연구 분야는 행동 내분비학이라고도 하며, 호르몬과 행동 간의 상호 작용에 대한 과학적 연구입니다. 호르몬은 행동에 영향을 미칠 수 있고 행동은 때때로 호르몬 농도에 영향을 미칠 수 있는 양방향 상호작용을 합니다. 호르몬은 내분비선에서 분비되어 혈액계를 통해 신경계에 영향을 주어 공격성, 짝짓기, 양육과 같은 행동을 조절하는 화학적 메신저입니다.
학습 목표
- Define the basic terminology and basic principles of hormone–behavior interactions.
- Explain the role of hormones in behavioral sex differentiation.
- Explain the role of hormones in aggressive behavior.
- Explain the role of hormones in parental behavior.
- Provide examples of some common hormone–behavior interactions.
- 호르몬-행동 상호작용의 기본 용어와 기본 원리를 정의합니다.
- 행동 성 분화에서 호르몬의 역할을 설명합니다.
- 공격적인 행동에서 호르몬의 역할을 설명합니다.
- 부모 행동에서 호르몬의 역할을 설명합니다.
- 몇 가지 일반적인 호르몬-행동 상호작용의 예를 제시합니다.
서론
이 모듈에서는 호르몬과 행동의 관계에 대해 설명합니다. 많은 독자는 호르몬이 행동에 영향을 미칠 수 있다는 일반적인 생각에 이미 익숙할 것입니다. 학생들은 일반적으로 혈중 성호르몬 농도가 사춘기 동안 증가하고 나이가 들어감에 따라, 특히 50세 이후에는 감소한다는 사실을 잘 알고 있습니다. 성적 행동도 비슷한 패턴을 보입니다. 또한 대부분의 사람들은 공격성과 단백 동화 스테로이드 호르몬의 관계에 대해 알고 있으며, 인공 스테로이드 호르몬을 투여하면 때때로 "로이드 레이지"라고 불리는 통제할 수 없는 폭력적인 행동이 발생한다는 사실도 알고 있습니다. 다양한 호르몬이 여러 유형의 행동에 영향을 미칠 수 있지만, 이 모듈에서는 호르몬과 행동의 몇 가지 예로만 한정하여 논의하겠습니다. 예를 들어, 행동의 성별 차이는 호르몬, 환경 또는 여러 요인의 조합의 결과일까요? 남성이 여성보다 공격적인 행동을 할 가능성이 훨씬 더 높은 이유는 무엇인가요? 호르몬이 소위 모성 '본능'을 매개하는 데 관여할까요? 행동 내분비학자들은 호르몬의 일반적인 생리적 효과가 행동의 발달과 표현을 어떻게 변화시키는지, 그리고 행동이 호르몬의 효과에 어떻게 영향을 미칠 수 있는지에 관심을 갖고 있습니다. 이 모듈에서는 호르몬이 행동에 미치는 영향을 현상학적으로나 기능적으로 설명합니다.
호르몬과 행동의 관계를 이해하려면 호르몬에 대해 간략히 설명하는 것이 중요합니다. 호르몬은 내분비선이라고 하는 특수 분비선에서 생성 및 방출되는 유기 화학 물질입니다. 호르몬은 이러한 분비샘에서 혈액으로 방출되어 발원지에서 어느 정도 떨어진 표적 구조에 작용하기 위해 이동할 수 있습니다. 호르몬은 신경계에서 동물의 활동을 조정하는 데 사용하는 화학 물질인 신경 전달 물질과 기능이 유사합니다. 그러나 호르몬은 신경전달물질보다 더 먼 거리와 훨씬 더 넓은 시간적 범위에서 작용할 수 있습니다(중점 주제 1). 행동에 영향을 미치는 호르몬의 예로는 테스토스테론(일반적인 안드로겐 유형), 에스트라디올(일반적인 에스트로겐 유형), 프로게스테론(일반적인 프로게스틴 유형), 코티솔(일반적인 글루코코르티코이드 유형)과 같은 스테로이드 호르몬이 있습니다(표 1, A-B). 옥시토신, 바소프레신, 프로락틴, 렙틴 등 여러 종류의 단백질 또는 펩타이드(작은 단백질) 호르몬도 행동에 영향을 미칩니다.
신경 전달과 호르몬 통신의 차이점
신경과 호르몬의 소통은 모두 화학적 신호에 의존하지만, 몇 가지 두드러진 차이점이 존재합니다. 신경계의 의사소통은 기차를 타고 여행하는 것과 비슷합니다. 출발지와 목적지 사이에 선로가 존재한다면 여행 계획에서 기차를 이용할 수 있습니다. 마찬가지로 신경 메시지는 기존 신경로를 따라 목적지까지만 이동할 수 있습니다. 반면에 호르몬 통신은 자동차를 타고 여행하는 것과 같습니다. 철로보다 도로가 더 많기 때문에 기차 여행보다 더 많은 목적지까지 운전할 수 있습니다. 마찬가지로 호르몬 메시지는 순환계를 통해 신체 어디든 이동할 수 있으며, 혈액을 받는 모든 세포는 잠재적으로 호르몬 메시지를 수신할 수 있습니다.
신경과 호르몬의 통신은 다른 방식으로도 다릅니다. 이를 설명하기 위해 디지털 기술과 아날로그 기술의 차이점을 생각해 보세요. 신경 메시지는 디지털 방식으로 발생과 소멸이 빠른 이벤트이며, 신경 신호는 밀리초 단위로 발생할 수 있습니다. 따라서 신경계는 상대적으로 빠른 신체의 변화를 매개합니다. 예를 들어, 신경계는 즉각적인 음식 섭취를 조절하고 신체 움직임을 지시합니다. 이와는 대조적으로 호르몬 메시지는 몇 초, 몇 분, 심지어 몇 시간이 걸릴 수 있는 아날로그적이고 단계적인 사건입니다. 호르몬은 성장, 발달, 생식, 신진대사와 같은 장기적인 과정을 매개할 수 있습니다.
호르몬과 신경 메시지는 모두 본질적으로 화학 물질이며 세포에서 비슷한 방식으로 방출되고 수신되지만, 중요한 차이점도 있습니다. 뉴런이 사용하는 화학적 메신저인 신경전달물질은 시냅스 후 뉴런의 막까지 20~30나노미터(30 X 10-9m)에 불과한 거리를 이동하여 수용체와 결합합니다. 호르몬은 순환계로 들어가서 특정 수용체와 결합하는 표적 세포에 도달하기까지 1밀리미터에서 2미터 이상 이동할 수 있습니다.
신경 통신과 호르몬 통신의 또 다른 차이점은 기능을 자발적으로 제어할 수 있는 정도입니다. 일반적으로 호르몬 신호보다 신경 신호에 대한 자발적 제어가 더 많습니다. 예를 들어 갑상선 호르몬 수치의 변화를 의지하는 것은 사실상 불가능한 반면, 명령에 따라 팔다리를 움직이는 것은 쉽습니다.
이러한 중요한 차이점에도 불구하고 신경계와 내분비계의 구분은 신경계가 호르몬 통신을 조절하는 방법에 대해 더 많이 알게 되면서 더욱 모호해지고 있습니다. 신경내분비학이라고 불리는 내분비계와 신경계 사이의 인터페이스를 더 잘 이해하면 호르몬과 행동 간의 상호 작용에 대한 향후 연구에서 중요한 진전을 이룰 수 있을 것입니다.


호르몬은 신체 기능을 조절, 통합 및 제어하여 개인의 생리와 행동을 조정합니다. 진화 과정에서 호르몬은 종종 신경계에 의해 생식 성공을 보장하기 위해 행동에 영향을 미치도록 선택되어 왔습니다. 예를 들어 생식세포(난자 또는 정자)의 성숙을 유발하는 테스토스테론과 에스트라디올이라는 동일한 호르몬은 짝짓기 행동도 촉진합니다. 이러한 이중 호르몬 기능은 동물이 수정에 사용할 수 있는 성숙한 생식세포를 가지고 있을 때 짝짓기 행동이 일어나도록 보장합니다. 생리적 및 행동 기능에 대한 내분비 조절의 또 다른 예는 임신에 의해 제공됩니다. 임신 중에는 에스트로겐과 프로게스테론 농도가 상승하며, 이러한 호르몬은 종종 어미의 모성 행동을 매개하는 데 관여합니다.
모든 세포가 모든 호르몬의 영향을 받는 것은 아닙니다. 오히려 특정 호르몬은 해당 호르몬에 대한 특정 호르몬 수용체를 가진 세포에만 직접적인 영향을 미칠 수 있습니다. 이러한 특정 수용체를 가진 세포를 호르몬의 표적 세포라고 합니다. 호르몬과 수용체의 상호 작용은 결국 효소 경로의 활성화로 이어지거나 단백질 합성을 조절하는 유전자 활성화를 켜거나 끄는 일련의 세포 이벤트를 시작합니다. 새로 합성된 단백질은 다른 유전자를 활성화하거나 비활성화하여 또 다른 일련의 세포 이벤트를 일으킬 수 있습니다. 중요한 것은 특정 호르몬이 효과를 발휘하려면 충분한 수의 적절한 호르몬 수용체가 있어야 한다는 것입니다. 예를 들어 테스토스테론은 남성의 성적 행동에 중요합니다. 남성의 테스토스테론이 너무 적으면 성적 동기가 낮아질 수 있으며, 테스토스테론 치료를 통해 회복할 수 있습니다. 그러나 남성의 테스토스테론 수치가 정상이거나 심지어 높은데도 성욕이 낮다면 수용체 부족이 원인일 수 있으며 추가 호르몬 치료가 효과적이지 않을 수 있습니다.
호르몬이 행동에 어떤 영향을 미칠 수 있나요? 인간과 다른 동물의 행동은 개념적으로 세 가지 상호 작용하는 요소로 구성되어 있다고 생각할 수 있습니다: (1) 입력 시스템(감각 시스템), (2) 통합자(중추 신경계), (3) 출력 시스템 또는 이펙터(예: 근육)가 그것입니다. 호르몬은 행동 변화를 일으키지 않습니다. 오히려 호르몬은 이 세 가지 시스템에 영향을 미쳐 특정 자극이 적절한 행동 또는 사회적 맥락에서 특정 반응을 유도할 가능성이 높아집니다. 즉, 호르몬은 적절한 상황에서 특정 행동이 나타날 확률을 변화시킵니다(Nelson, 2011). 이는 호르몬과 행동의 관계를 생각하는 방식에 영향을 미칠 수 있는 중요한 구분입니다.
이 세 가지 요소로 구성된 행동 체계를 얼룩부리새의 노래라는 단순한 행동에 적용할 수 있습니다. 수컷 제브라 핀치만이 노래를 부릅니다. 성체 수컷 핀치새의 고환을 제거하면 새들은 노래를 줄이지만, 거세한 핀치새는 고환을 다시 이식하거나 테스토스테론 또는 에스트라디올로 치료하면 노래를 다시 시작합니다. 일반적으로 안드로겐은 "남성" 호르몬이고 에스트로겐은 "여성" 호르몬이라고 생각하지만, 테스토스테론은 신경 세포에서 에스트라디올로 전환되는 것이 일반적입니다(그림 1). 따라서 많은 남성다운 행동은 에스트로겐의 작용과 관련이 있습니다! 실제로 모든 에스트로겐은 일반적인 생화학적 합성 과정으로 인해 먼저 안드로겐에서 전환되어야 합니다. 전환 효소가 부족하거나 결핍되면 여성은 안드로겐을 과도하게 생산하여 관련 남성 형질을 개발할 수 있습니다. 또한 환경의 에스트로겐이 사람을 포함한 동물의 신경계에 영향을 미칠 수도 있습니다(예: Kidd et al., 2007). 다시 말하지만, 노래하는 행동은 혈중 테스토스테론 또는 에스트로겐 농도가 높을 때 가장 빈번하게 나타납니다. 수컷은 짝을 유인하거나 잠재적 경쟁자를 영역에서 쫓아내기 위해 노래를 부릅니다.
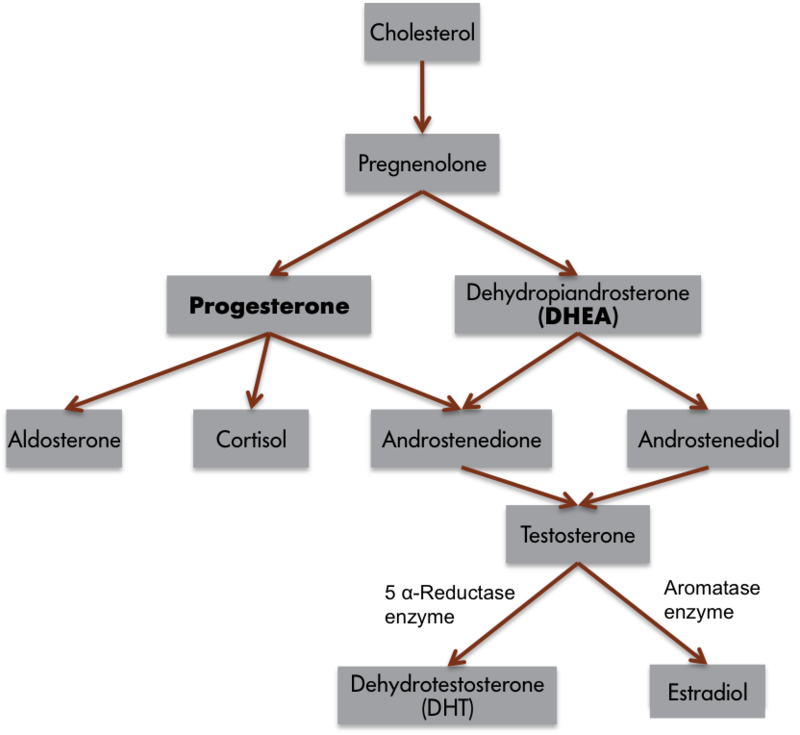
이러한 관찰을 통해 에스트로겐이 노래에 어떤 식으로든 관여한다는 것이 분명해졌지만, 방금 소개한 세 가지 구성 요소 프레임워크가 이 행동에서 에스트로겐의 역할을 탐구하기 위한 가설을 세우는 데 어떻게 도움이 될 수 있을까요? 입력 시스템을 조사함으로써 에스트로겐이 새의 감각 능력을 변화시켜 일반적으로 노래를 유도하는 환경 단서를 더 두드러지게 만드는지 여부를 확인할 수 있습니다. 만약 그렇다면 암컷이나 경쟁자를 더 쉽게 보거나 들을 수 있을 것입니다. 에스트로겐은 중추 신경계에도 영향을 미칠 수 있습니다. 에스트로겐이 존재하면 신경 구조나 신경 처리 속도가 달라질 수 있습니다. 더 높은 신경 과정(예: 동기 부여, 주의력 또는 지각)도 영향을 받을 수 있습니다. 마지막으로, 에스트로겐의 영향을 받는 기관인 근육은 에스트로겐의 존재에 의해 영향을 받을 수 있습니다. 혈중 에스트로겐 농도는 새의 발성 기관인 송새의 시린스 근육에 어떤 식으로든 영향을 미칠 수 있습니다. 따라서 에스트로겐은 개별 새의 감각 능력, 중추 처리 시스템 또는 이펙터 기관에 영향을 미쳐 새소리에 영향을 미칠 수 있습니다. 테스토스테론에서 유래한 에스트로겐이 새소리에 어떤 영향을 미치는지 완전히 이해하지는 못했지만, 대부분의 경우 호르몬은 이 세 가지 요소 중 하나, 둘 또는 모두에 영향을 미쳐 행동에 영향을 미치는 것으로 간주할 수 있으며, 이 세 가지 프레임워크는 이러한 문제를 탐구하기 위한 가설과 실험을 설계하는 데 도움이 될 수 있습니다.
행동이 호르몬에 어떤 영향을 미칠 수 있나요? 새소리의 예는 호르몬이 행동에 어떻게 영향을 미칠 수 있는지를 보여 주지만, 앞서 언급했듯이 상호 관계, 즉 행동이 호르몬 농도에 영향을 미칠 수 있다는 사실도 알 수 있습니다. 예를 들어, 영역 침입자를 목격하면 서식하는 수컷 새의 혈중 테스토스테론 농도가 높아져 노래하거나 싸우는 행동이 자극될 수 있습니다. 마찬가지로, 싸움에서 패배한 수컷 쥐나 붉은털원숭이는 그 후 며칠 또는 몇 주 동안 순환하는 테스토스테론 농도가 감소합니다. 인간에게도 비슷한 결과가 보고되었습니다. 테스토스테론 농도는 실제 전투에 참여하는 사람뿐만 아니라 모의 전투에 참여하는 사람에게도 영향을 미칩니다. 예를 들어, 지역 체스 토너먼트에서 승자는 테스토스테론 농도가 높아지고 패자는 테스토스테론 농도가 감소했습니다.

경기 결과에 따라 호르몬이 영향을 받기 위해 경기에 직접 참여하지 않아도 됩니다. 1994년 월드컵 축구 결승전 전후에 테스토스테론 분석을 위해 브라질과 이탈리아 팀의 남성 팬을 모집하여 타액 샘플을 제공했습니다. 브라질과 이탈리아는 결승전까지 동점을 기록했지만 마지막 순간 브라질이 페널티킥으로 승리했습니다. 브라질 팬들은 환호했고 이탈리아 팬들은 상심에 빠졌습니다. 샘플을 분석했을 때, 경기 전 기준치에 비해 샘플을 채취한 12명의 브라질 팬 중 11명은 테스토스테론 농도가 증가했고, 9명의 이탈리아 팬 중 9명은 테스토스테론 농도가 감소했습니다(Dabbs, 2000).
어떤 경우에는 호르몬이 행동에 대한 기대감에 의해 영향을 받을 수 있습니다. 예를 들어 테스토스테론 농도는 여성의 성적 동기와 행동에도 영향을 미칩니다. 한 연구에서는 성관계와 테스토스테론의 상호작용을 여성의 다른 활동(껴안기 또는 운동)과 비교했습니다(van Anders, Hamilton, Schmidt, & Watson, 2007). 여성들은 세 차례에 걸쳐 활동 전, 활동 후, 다음 날 아침 타액 샘플을 제공했습니다. 분석 결과, 여성의 테스토스테론은 다른 시간에 비해 성관계 전에 상승하는 것으로 확인되었습니다. 따라서 성적 행동과 테스토스테론 사이에는 기대 관계가 존재합니다. 테스토스테론 수치는 운동에 비해 성관계 후 더 높았는데, 이는 성행위가 여성의 호르몬 농도에 영향을 미칠 수 있음을 시사합니다.
성별 차이
암탉과 수탉은 다릅니다. 소와 황소는 다릅니다. 남자와 여자는 다릅니다. 소녀와 소년도 다릅니다. 인간은 많은 동물과 마찬가지로 신체의 크기와 모양, 생리학, 그리고 우리의 목적에 따라 행동에 있어 성적 이형성(di, "둘", morph, "유형")을 가지고 있습니다. 남학생과 여학생의 행동은 여러 가지 면에서 다릅니다. 일반적으로 여자아이는 남자아이에 비해 언어 능력이 뛰어나며, 남자아이는 난독증(읽기 장애)과 말더듬증에 걸릴 확률이 여자아이보다 거의 2배, 자폐증에 걸릴 확률은 거의 4배나 높습니다. 일반적으로 남자아이가 여자아이보다 시공간 능력이 필요한 작업에서 더 우수합니다. 여자아이는 남자아이보다 양육 행동에 더 자주 참여합니다. 모든 신경성 식욕 부진 사례의 90% 이상이 젊은 여성과 관련이 있습니다. 젊은 남성은 젊은 여성보다 조현병에 걸릴 확률이 두 배나 높습니다. 남자아이는 여자아이보다 훨씬 더 공격적이며 일반적으로 거칠고 거친 놀이에 더 많이 참여합니다(Berenbaum, Martin, Hanish, Briggs, & Fabes, 2008). 공격성의 차이와 같은 많은 성별 차이는 성인이 되어서도 지속됩니다. 예를 들어, 폭력적인 행동으로 징역형을 선고받는 남성이 여성보다 훨씬 더 많습니다. 남성과 여성의 호르몬 차이는 사춘기에 나타나는 성인 성차를 설명할 수 있지만, 사춘기 이전의 아동과 생식선 활성화 이전의 행동 성차를 설명하는 것은 무엇일까요? 발달하는 생식선에서 분비되는 호르몬은 개인이 남성 또는 여성으로 발달할지 여부를 결정합니다. 포유류의 배아 고환은 펩타이드 호르몬뿐만 아니라 안드로겐을 생성하여 신체, 중추 신경계 및 후속 행동의 발달을 남성적인 방향으로 유도합니다. 포유류의 배아 난소는 사실상 정지 상태이며 고농도의 호르몬을 분비하지 않습니다. 난소가 있거나 생식선이 전혀 없는 경우, 형태적, 신경적, 그리고 나중에 행동 발달은 여성의 경로를 따릅니다.

생식선 스테로이드 호르몬은 뇌와 행동에 조직적(또는 프로그래밍) 영향을 미칩니다(Phoenix, Goy, Gerall, & Young, 1959). 스테로이드 호르몬의 조직화 효과는 발달 초기 단계에 상대적으로 제한적입니다. 포유류의 행동 조직화에 대한 고환과 난소의 영향에는 비대칭성이 존재합니다. 생후 초기의 호르몬 노출은 이후 설치류의 행동에 조직적인 영향을 미치며, 초기 스테로이드 호르몬 치료는 설치류의 행동(짝짓기 및 공격성)을 비교적 비가역적이고 영구적으로 남성화합니다. 이러한 초기 호르몬 효과는 성인기에 제공되는 스테로이드 호르몬의 가역적 행동 영향과 대조될 수 있으며, 이를 활성화 효과라고 합니다. 성인 행동에 대한 호르몬의 활성화 효과는 일시적이며 호르몬이 대사된 후 곧 사라질 수 있습니다. 따라서 전형적인 남성 행동은 임신 중(인간의 경우) 또는 출생 직후(설치류의 경우) 안드로겐에 노출되어 뇌를 어느 정도 남성화해야 하며, 사춘기 중 또는 사춘기 이후에 이러한 신경 회로를 활성화하기 위해 안드로겐이 필요합니다. 전형적인 여성 행동은 어릴 때 안드로겐에 노출되지 않아 뇌가 여성화되고, 성인이 되어 이러한 신경 회로를 활성화하려면 에스트로겐이 필요합니다. 하지만 이 단순한 이분법은 행동에 있어 매우 뚜렷한 성적 이형성을 가진 동물에게 잘 작동하지만 사람에게 적용할 때는 많은 주의가 필요합니다.
대형 장난감 가게에 가보면 분홍색 상자로 가득 찬 통로 몇 개와 인접한 통로에는 분홍색 장난감 포장이 전혀 없는 것을 볼 수 있을 것입니다. 놀랍게도 이러한 통로에서 남학생과 여학생이 강하게 분리되어 있는 것을 볼 수 있습니다. "분홍색"통로에서 소년을 보는 것은 드물고 그 반대의 경우도 마찬가지입니다. 장난감 제조업체는 종종 성 편향적인 장난감을 만든다는 비난을 받기도 하지만, 남자아이와 여자아이가 특정 유형과 색상의 장난감을 가지고 노는 것을 더 좋아하는 것 같습니다. 실제로 장난감 제조업체는 남녀 모두에게 장난감을 판매할 수 있다면 매출이 즉시 두 배로 늘어날 것입니다. 일반적으로 남자아이는 트럭이나 공과 같은 장난감을 선호하고 여자아이는 인형과 같은 장난감을 선호합니다. Y 염색체에 장난감 자동차와 트럭에 대한 선호도를 암호화하는 유전자가 있는지는 확실하지 않지만, 호르몬이 특정 유형의 장난감이나 놀이 행동 스타일을 선호하도록 아동의 뇌 발달을 형성할 수 있습니다. 아이들은 성별에 따라 어떤 유형의 장난감과 어떤 놀이 스타일이 적합한지 학습한다고 보는 것이 합리적입니다. 그렇다면 인간 행동의 성별 차이를 이해하기 위한 학습에서 생리적 메커니즘의 기여를 어떻게 이해하고 분리할 수 있을까요? 이러한 문제를 풀기 위해 동물 모델이 자주 사용됩니다. 성별 차이가 보통 정도의 문제일 뿐인 인간의 상황과 달리, 일부 동물의 경우 한 성별의 구성원만 특정 행동을 보일 수 있습니다. 앞서 언급했듯이 수컷 송버드만 노래하는 경우가 많습니다. 이처럼 성에 따라 강하게 편향된 행동에 대한 연구는 행동, 호르몬, 신경계 간의 상호작용을 이해하는 데 특히 유용합니다.
버벳 원숭이에 대한 한 연구는 장난감 선호도 설정에 있어 학습의 우선순위에 의문을 제기합니다(Alexander & Hines, 2002). 암컷 버베트 원숭이는 인형이나 냄비 같은 여아형 장난감을 선호한 반면, 수컷 버베트 원숭이는 자동차나 공 같은 남아형 장난감을 선호했습니다. 그림책이나 인형과 같은 성 중립적인 장난감에 대한 선호도에는 성별 차이가 없었습니다. 아마도 원숭이에게는 '남자' 또는 '여자' 장난감에 대한 사전 개념이 없는 것으로 추정됩니다. 어린 붉은털원숭이도 비슷한 장난감 선호도를 보입니다.

그렇다면 장난감 선호도의 성별 차이의 근간은 무엇일까요? 장난감(또는 물건)의 특정 속성이 남아 또는 여아에게 어필할 수 있습니다. 남아 또는 수컷 붉은털원숭이 또는 붉은털원숭이에게 어필하는 장난감은 공이나 장난감 자동차처럼 공간에서 활발하게 움직일 수 있는 물체, 활동적이고 거칠고 넘어지는 놀이에 포함될 수 있는 장난감입니다. 여아 또는 암컷 버벳원숭이가 선호하는 장난감의 매력은 색상에 기반한 것으로 보입니다. 분홍색과 빨간색(인형과 냄비의 색상)은 유아의 주의를 끌 수 있습니다.
사회는 성별에 따른 장난감에 대한 이러한 고정관념적 반응을 강화할 수 있습니다. 장난감 선호도의 성별 차이는 생후 12개월 또는 24개월에 나타나고 생후 36개월이 되면 고정된 것처럼 보이지만, 생후 1년 동안에도 장난감 선호도의 성별 차이가 존재하나요? 말을 하기 전의 영아에게 무엇을 선호하는지 물어보기는 어렵지만, 연구자들이 아기가 여러 장난감을 보는 시간을 조사한 연구에서 시선추적 데이터에 따르면 3개월의 영아는 장난감 선호도에 성별 차이가 있었으며, 여아는 인형을 선호한 반면 남아는 트럭을 선호했습니다. 호르몬이 장난감 선호도에 영향을 미친다는 것을 증명하지는 못하지만 암시하는 또 다른 결과는 선천성 부신 과형성증(CAH) 진단을 받은 여아 중 부신에서 다양한 양의 안드로겐이 생성되는 여아가 그렇지 않은 여아보다 남성적인 장난감을 더 자주 가지고 노는 것을 관찰한 것입니다. 또한 장애의 정도(즉, 태아기 안드로겐 노출 정도)와 놀이 행동의 남성화 정도 간에 용량 반응 관계가 관찰되었습니다. 예를 들어 장난감 선호도나 놀이 활동의 성별 차이는 남아와 여아의 내분비 환경 차이로 인한 불가피한 결과일까요, 아니면 문화적 관습과 신념에 의해 부과된 차이일까요? 이러한 차이는 어릴 때부터 성별에 특화된 장난감을 받은 결과일까요, 아니면 내분비적 요인과 문화적 요인이 복합적으로 작용한 결과일까요? 다시 말하지만, 이러한 질문은 사람들에게 풀기 어려운 질문입니다.
행동의 성별 차이가 발달 초기에 나타나더라도 사회적 기대의 영향에 대해서는 의문이 있는 것 같습니다. 한 가지 예로 수컷이 더 신체적인 놀이를 하는 인간의 놀이 행동 패턴을 들 수 있는데, 이러한 패턴은 인간이 아닌 영장류, 쥐, 개 등 다른 여러 종에서도 볼 수 있습니다. 남자아이와 여자아이의 거친 놀이 빈도의 차이는 남성 또는 여성이라는 생물학적 요인 때문일까요, 아니면 문화적 기대와 학습 때문일까요? 생물학적 영향과 문화적 영향이 서로 복합적으로 작용하여 거친 놀이의 빈도를 조절한다면, 남녀 간 차이의 몇 퍼센트가 생물학적 요인에 기인하고 몇 퍼센트가 사회적 영향에 기인하는 것일까요? 중요한 것은 이러한 특성이 사실상 항상 불연속적인 범주가 아닌 연속체를 따라 배열될 때 '정상적인' 성차에 대해 이야기하는 것이 적절할까요?
성 차이는 인간과 비인간 동물에게 공통적으로 존재합니다. 남성과 여성은 안드로겐과 에스트로겐 스테로이드 호르몬 농도의 비율이 다르기 때문에 행동 내분비학자들은 특히 호르몬에 의해 행동의 성차가 매개되는 정도에 관심을 가져왔습니다. 여성 또는 남성이 되는 과정을 성 분화라고 합니다. 성 분화의 주요 단계는 수정에서 발생합니다. 포유류의 경우, 난자(항상 X 염색체를 포함하고 있음)는 Y 또는 X 염색체를 가진 정자에 의해 수정될 수 있으며, 이 과정을 성 결정이라고 합니다. 동형 접합 포유류(XX)의 염색체 성은 암컷이고, 이형 접합 포유류(XY)의 염색체 성은 수컷입니다. 염색체 성별은 생식선 성별을 결정합니다. 사실상 모든 후속 성적 분화는 일반적으로 생식선 스테로이드 호르몬에 대한 차등 노출의 결과입니다. 따라서 생식선 성은 호르몬 성을 결정하며, 호르몬 성은 형태학적 성을 조절합니다. 중추 신경계와 근육과 같은 일부 이펙터 기관의 형태 학적 차이는 행동 성별 차이로 이어집니다. 성적 분화 과정은 복잡하며 오류의 가능성이 있습니다. 안드로겐에 대한 주 산기 노출은 여성들 사이에서 비정상적인 성적 분화의 가장 흔한 원인입니다. 안드로겐의 공급원은 내부(예: 부신에서 분비되는 것) 또는 외부(예: 환경 에스트로겐에 노출되는 것)일 수 있습니다. 터너 증후군은 두 번째 X 염색체가 없거나 손상되었을 때 발생하며, 이러한 사람들은 이형성 난소를 가지고 있고 사춘기까지 스테로이드 호르몬에 노출되지 않습니다. 흥미롭게도 터너 증후군을 가진 여성은 종종 공간 기억력이 손상됩니다.
암컷 포유류는 "중성" 성으로 간주되며, 수컷 분화에는 추가적인 생리적 단계가 필요하며 단계가 많을수록 분화 오류의 가능성이 높아집니다. 남성의 비정상적인 성 분화의 몇 가지 예로는 5α-환원효소 결핍증(디하이드로테스토스테론이 부족하여 모호한 생식기를 가지고 태어나 암컷으로 자랐지만 사춘기에 남성화가 일어나는 경우)과 안드로겐 무감각 증후군 또는 TFM(안드로겐 수용체가 부족하여 암컷으로 발달하는 경우)이 있습니다. 행동 내분비학자들은 여성 또는 남성이라는 이분법적 범주에 깔끔하게 속하지 않고 성 분화 과정이 비정형적인 개인을 연구함으로써 전형적인 성 분화 과정에 대한 힌트를 얻습니다.
궁극적으로 호르몬이 인간의 뇌와 행동에서 성 차이를 매개하는 방법(이러한 차이가 어느 정도까지 발생하는지)을 알고자 할 수 있습니다. 뇌와 행동의 성별 차이의 근본적인 메커니즘을 이해하기 위해 새소리의 예로 돌아가 보겠습니다. 새는 행동의 성별 차이가 호르몬에 의해 유도된 뇌의 구조적 변화의 결과라는 가장 좋은 증거를 제공합니다(Goodson, Saldanha, Hahn, & Soma, 2005). 신경 조직의 구조적 차이가 행동과 직접적으로 연결되지 않는 포유류와 달리 조류 뇌의 구조적 차이는 새소리라는 성 행동과 직접적으로 연결되었습니다.
송새의 여러 뇌 영역은 크기에서 성별에 따라 상당한 차이를 보입니다. (1) 노래 생성 운동 경로와 (2) 청각 전달 경로라는 두 가지 주요 뇌 회로 경로가 새소리의 학습과 생성에 관여하는 것으로 밝혀졌습니다. 수컷 제브라 핀치의 노래 생성 경로의 일부 영역은 암컷보다 3~6배 더 큽니다. 이 뇌 영역의 크기가 더 크다는 것은 이 핵의 뉴런이 더 크고, 더 많고, 더 멀리 떨어져 있다는 것을 반영합니다. 성체 수컷 새를 거세하면 노래가 줄어들지만, 노래 생성을 조절하는 뇌핵의 크기는 줄어들지 않습니다. 마찬가지로, 성체 암컷 제브라 핀치의 안드로겐 치료는 노래 또는 노래 제어 영역의 크기 변화를 유도하지 않습니다. 따라서 스테로이드 호르몬의 활성화 효과는 제브라 핀치의 노래 행동이나 뇌핵 크기의 성별 차이를 설명하지 못합니다. 이러한 구조의 성 차이는 에스트라 디올 (남성화) 또는 스테로이드 부족 (여성화)에 의해 난자에서 조직되거나 프로그래밍됩니다.
종합하면, 에스트로겐은 새의 노래 시스템의 기초가 되는 신경 메커니즘을 활성화하는 데 필요한 것으로 보입니다. 새의 고환은 주로 안드로겐을 생산하며, 이 안드로겐은 순환계로 들어갑니다. 안드로겐은 아로마타제를 함유한 뉴런으로 들어가 에스트로겐으로 전환됩니다. 실제로 뇌는 많은 조류 종에서 남성적인 행동을 활성화하는 에스트로겐의 주요 공급원입니다.
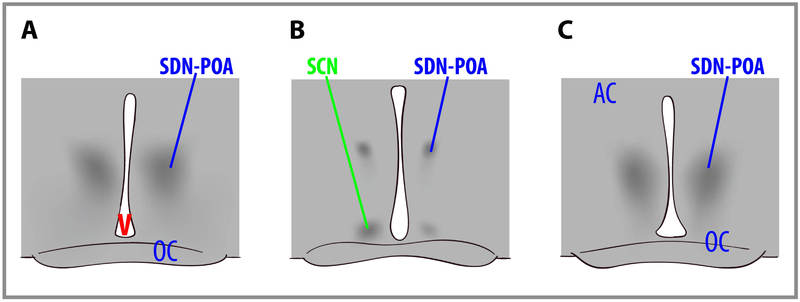
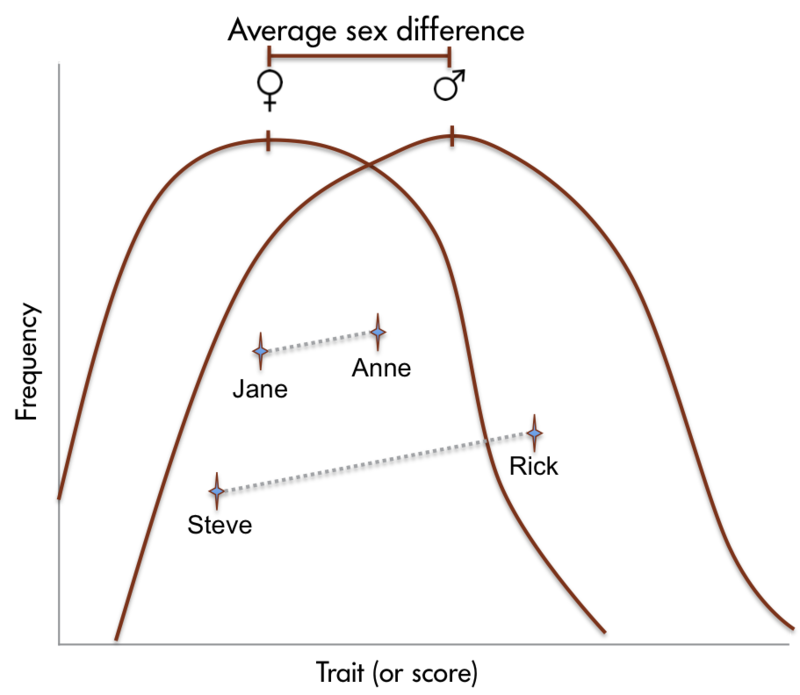
인간의 뇌 크기에서 성별 차이가 있다는 사실은 수년 동안 보고되어 왔습니다. 최근에는 특정 뇌 구조에서 성별 차이가 발견되었습니다(그림 2). 여러 인지 기능의 성별 차이도 보고되었습니다. 일반적으로 여성은 청각 정보에 더 민감한 반면, 남성은 시각 정보에 더 민감합니다. 또한 여성은 일반적으로 미각과 후각 입력에 남성보다 더 민감합니다. 여성은 남성보다 인지 기능의 측면화가 덜 나타납니다. 평균적으로 여성은 일반적으로 언어, 지각, 소근육 운동 능력이 뛰어난 반면, 남성은 지도 읽기, 방향 찾기 등 양적, 시각적 공간 작업에서 여성보다 뛰어납니다. 신뢰할 수 있는 성별 차이가 문서화되어 있기는 하지만, 이러한 능력 차이는 미미합니다. 대부분의 인지 능력에서 성별 간 차이보다 성별 내 차이가 더 크다는 점에 유의해야 합니다(그림 3).
공격적인 행동
공격적인 행동의 가능성은 두 명 이상의 이해관계가 충돌할 때마다 존재합니다(Nelson, 2006). 갈등은 영역, 먹이, 짝과 같은 제한된 자원을 놓고 발생할 가능성이 가장 높습니다. 사회적 상호작용에 따라 어떤 동물이 경쟁 자원에 접근할 수 있는지가 결정됩니다. 많은 경우, 한 동물이 복종하는 자세나 제스처를 취하면 자원을 둘러싼 실제 전투를 피할 수 있습니다. 동물은 우위를 결정하기 위한 위협 표시나 의식적인 전투에 참여할 수도 있지만 신체적 피해는 가하지 않습니다.
안드로겐 스테로이드 호르몬이 여러 종에 걸쳐 공격적인 행동을 매개한다는 압도적인 정황 증거가 있습니다. 첫째, 테스토스테론의 혈장 농도의 계절적 변화와 공격성의 계절적 변화가 일치합니다. 예를 들어, 수컷 사슴은 테스토스테론이 많이 분비되는 가을에 공격적인 행동의 발생률이 최고조에 이릅니다. 둘째, 고환이 활성화되고 안드로겐의 혈중 농도가 상승하는 사춘기에는 공격적인 행동이 증가합니다. 어린 사슴은 짝짓기 시즌에는 싸움에 참여하지 않습니다. 셋째, 특정 종에서 수컷은 일반적으로 암컷보다 더 공격적입니다. 사슴도 마찬가지입니다. 사슴에 비해 암사슴은 공격적인 행동을 거의 보이지 않으며, 드물게 나타나는 암사슴의 공격적인 행동은 공격적인 수컷의 공격적인 행동과는 질적으로 다릅니다. 마지막으로, 거세는 일반적으로 수컷의 공격성을 감소시키며 테스토스테론 대체 요법은 공격성을 거세 전 수준으로 회복시킵니다. 이 모듈의 범위를 벗어난 이러한 일반적인 관찰에는 몇 가지 흥미로운 예외가 있습니다.
앞서 언급했듯이 수컷은 일반적으로 암컷보다 더 공격적입니다. 물론 인간 수컷은 암컷보다 훨씬 더 공격적입니다. 북미에서는 여성보다 남성이 폭력 범죄로 더 많이 유죄 판결을 받습니다. 인간의 공격성에 대한 성별 차이는 매우 일찍 나타납니다. 학창 시절의 모든 연령대에서 여학생보다 남학생이 더 많은 신체적 폭행을 시작합니다. 거의 모든 사람들이 이러한 성별 차이가 존재한다는 것을 인정하지만, 인간의 행동에 성차라는 원인을 부여하는 것은 항상 많은 논쟁을 불러일으킵니다. 안드로겐이 공격적인 행동을 촉진하고 남자아이가 여자아이보다 혈중 안드로겐 농도가 높기 때문에 남자아이가 여자아이보다 더 공격적일 수 있습니다. 남아의 뇌가 태아기에 안드로겐에 노출되어 뇌의 '배선'이 공격성 표현을 용이하게 하는 방식으로 구성되어 있기 때문에 남아와 여아의 공격성이 다를 수 있습니다. 또한 남자아이는 가족, 또래 또는 다른 사람들로부터 공격적인 행동을 하도록 격려받고 여자아이는 낙담할 수도 있습니다. 이 세 가지 가설은 상호 배타적인 것은 아니지만, 인간의 공격성에 대한 성별 차이를 설명하기 위해 이 가설들을 구분하는 것은 매우 어렵습니다.

이러한 가설을 평가하려면 어떤 종류의 연구가 필요할까요? 일반적으로 인간의 행동 발달에 대한 환경과 생리학의 영향을 분리하기는 어렵습니다. 예를 들어, 남자아이와 여자아이는 아주 어린 나이에 거칠고 넘어지는 놀이에서 차이가 나며, 이는 공격성에 대한 초기 생리적 영향을 시사합니다. 그러나 부모는 암수 자녀와 다르게 상호작용하며, 일반적으로 암컷보다 수컷 유아에게 더 거칠게 놀아 공격성의 성별 차이가 부분적으로 학습된 것임을 시사합니다. 이러한 부모의 상호작용 스타일 차이는 생후 첫 주에 분명하게 드러납니다. 이처럼 인간 행동에 영향을 미치는 요인이 복잡하기 때문에 환경적 영향이 비교적 일정하게 유지될 수 있는 비인간 동물에서 성 분화 행동에 대한 호르몬의 영향을 연구해 왔습니다. 이 과정을 실험적으로 쉽게 조작할 수 있도록 출생 후 성 분화가 일어나는 동물 모델이 자주 사용됩니다.
다시 말하지만, 적절한 동물 모델을 사용하면 위에서 제기한 질문을 해결할 수 있습니다: 공격성의 성별 차이는 수컷의 성인 혈중 안드로겐 농도가 암컷보다 높기 때문인가, 아니면 수컷의 뇌가 주산기 호르몬에 의해 다르게 조직되기 때문에 암컷보다 수컷이 더 공격적인가? 수컷이 일반적으로 암컷보다 공격적인 이유는 초기 혈중 안드로겐 농도와 현재 혈중 안드로겐 농도의 상호 작용 때문일까요? 수컷 쥐를 생후 6일 이전에 거세한 후 성체가 되어 프로피오네이트 테스토스테론으로 치료하면 수컷 쥐는 낮은 수준의 공격성을 보입니다. 마찬가지로 생후 6일 이전에 난소 절제술을 받았지만 성체가 되어 안드로겐을 투여받은 암컷은 수컷과 같은 수준의 공격성을 나타내지 않습니다. 주산기 생식선 절제술을 받은 수컷 또는 암컷에게 생후 6일 이전과 성체기에 테스토스테론을 투여하면 일반적인 수컷 마우스에서 관찰되는 것과 유사한 수준의 공격성이 나타납니다. 따라서 생쥐에서 수컷이 암컷보다 더 공격적으로 행동하는 성향은 출생 전후에 안드로겐에 의해 조직화되지만 사춘기 이후에는 안드로겐의 존재가 완전히 발현될 필요가 있습니다. 즉, 수컷 생쥐의 공격성은 안드로겐에 의해 조직화되고 활성화됩니다. 스테로이드 호르몬에 의한 뇌의 사전 조직화 없이 성인기에 테스토스테론에 노출되면 전형적인 남성 수준의 공격성을 유발하지 않습니다. 따라서 집쥐의 공격적 행동에 대한 호르몬 조절은 다른 설치류 종에서 이성애 수컷의 짝짓기 행동에 대한 호르몬 매개와 유사합니다. 공격적인 행동은 쥐, 햄스터, 들쥐, 개, 그리고 일부 영장류 종을 포함한 많은 종에서 안드로겐에 의해 조직화되고 활성화됩니다.
부모의 행동
부모의 행동은 암컷의 몸을 떠난 수정란이나 새끼의 생존에 직접적으로 기여하는 모든 행동으로 간주할 수 있습니다. 포유류의 부모 돌봄에는 여러 가지 패턴이 있습니다. 신생아의 발달 상태는 한 종에서 부모 돌봄의 유형과 질을 결정하는 중요한 요소입니다. 모성 돌봄은 부성 돌봄보다 훨씬 더 일반적입니다. 포유류의 부모 행동과 호르몬의 상관관계에 대한 대부분의 연구는 쥐를 대상으로 수행되었습니다. 쥐는 새끼를 낳으면 어미 쥐는 둥지를 짓고, 새끼 위에 웅크리고 앉아 수유를 하고, 따뜻함을 제공하고, 새끼를 찾고, 침입자를 향한 공격성을 높이는 등 정형화된 모성 행동을 합니다. 임신하지 않은 암컷 쥐(또는 수컷)를 새끼 쥐에게 노출시키면 가장 흔한 반응은 새끼 쥐로부터 멀리 웅크리는 것입니다. 쥐는 새로운 것을 피합니다(네오포비아). 그러나 성체 쥐를 매일 새끼 쥐에게 노출시키면 곧 모성애적인 행동을 하기 시작합니다. 이 과정을 오목화 또는 민감화라고 하며, 성체 쥐가 새끼 쥐에 대한 두려움을 줄이는 역할을 하는 것으로 보입니다.
물론 새로운 어미는 일주일이 아니라 새끼가 태어나자마자 모성 행동을 해야 합니다. 쥐의 모성 행동의 시작은 호르몬에 의해 매개됩니다. 호르몬 제거 및 대체 요법과 같은 여러 가지 연구 방법이 쥐의 모성 행동의 호르몬 상관 관계를 결정하는 데 사용되었습니다. 고농도의 에스트라디올과 아마도 프로락틴 및 옥시토신과 함께 고농도의 프로게스테론이 지속된 후 임신 말기에 혈중 프로게스테론 농도가 빠르게 감소하면 암컷 쥐는 새끼가 있는 상황에서 거의 즉시 모성 행동을 하도록 유도합니다. 분만 시 호르몬의 이러한 패턴은 새끼에 대한 성체 쥐의 일반적인 공포 반응을 무시하고 모성 행동의 시작을 허용합니다. 따라서 소위 모성 "본능"은 접근 경향을 높이고 회피 경향을 낮추기 위해 호르몬이 필요합니다. 실험용 생쥐와 쥐는 일반적으로 온순하지만, 어미는 새끼에게 너무 가까이 다가오는 동물에게 상당히 공격적일 수 있습니다. 프로게스테론은 설치류에서 이러한 모성 공격성을 유도하는 주요 호르몬으로 보이지만, 종에 따라 차이가 있습니다. 암컷의 행동에서 모성 공격성의 역할은 적절하게 설명되거나 테스트되지 않았습니다.

앨리슨 플레밍과 그녀의 공동 연구자들은 일련의 우아한 실험을 통해 인간 어머니의 행동과 내분비 상관관계, 그리고 자기보고 설문지에 나타난 모성 태도의 내분비 상관관계를 연구했습니다. 아기를 쓰다듬거나 안아주거나 입맞추는 등의 반응을 애정 행동이라고 하고, 아기에게 말하거나 노래하거나 달래는 행동을 음성 행동으로 간주했습니다. 애정적 행동과 음성적 행동 모두 접근 행동으로 간주했습니다. 기저귀 갈기, 트림시키기 등 기본적인 돌봄 활동도 기록했습니다. 이 연구에서 태도 설문지를 통해 측정한 호르몬 농도와 산모의 반응성 사이에는 아무런 관계가 발견되지 않았습니다. 예를 들어, 대부분의 여성은 임신 초기에 긍정적인 자아상을 보이다가 임신 후반기에 하락했지만 출산 후에는 회복되었습니다. 모성애와 관련된 감정의 하락은 임신 후반기에 발생했지만 대부분의 여성에서 출산 후 크게 반등했습니다. 그러나 설문 응답이 아닌 행동을 호르몬 농도와 비교했을 때 다른 이야기가 나타났습니다. 혈장 내 코르티솔 농도는 접근 행동과 양의 상관관계가 있었습니다. 즉, 수유 직전 또는 직후에 채취한 샘플에서 혈중 코르티솔 농도가 높은 여성은 코르티솔 농도가 낮은 어머니에 비해 아기에게 신체적으로 애정 어린 행동을 더 많이 하고 더 자주 말을 걸었습니다. 이 연구의 추가 분석 결과, 임신 기간 동안 긍정적인 모성애(감정과 태도)를 느꼈다고 보고한 산모의 경우 상관관계가 훨씬 더 큰 것으로 나타났습니다. 실제로 여성 간 모성 행동 변화의 거의 절반은 임신 중 코르티솔 농도와 긍정적인 모성 태도로 설명할 수 있습니다.
아마도 코르티솔은 산모의 행동을 직접적으로 유도하지는 않지만, 산모의 전반적인 각성 수준을 증가시켜 영아가 생성하는 신호에 대한 반응성을 증가시킴으로써 산모 돌봄의 질에 간접적으로 작용할 수 있습니다. 또한 코르티솔 농도가 높은 산모는 영아의 냄새에 더 끌리고, 영아를 식별하는 능력이 뛰어나며, 일반적으로 영아의 단서를 매우 매력적으로 여기는 것으로 나타났습니다(Fleming, Steiner, & Corter, 1997).
내측 전전두엽 영역은 쥐의 모성 행동 표현에 매우 중요합니다. 편도체는 모성 행동의 발현을 강압적으로 억제하는 것으로 보입니다. 성체 쥐는 새끼 쥐를 두려워하는데, 이러한 반응은 화학 감각 정보에 의해 매개되는 것으로 보입니다. 편도체의 병변 또는 보체 기관에서 편도체로의 구심성 감각 경로는 모성 행동의 발현을 억제합니다. 호르몬이나 감작이 편도체를 억제하는 작용을 하여 모성 행동의 발현을 허용할 가능성이 높습니다. 상관관계는 확립되었지만, 인간 산모의 뇌 구조적 변화에 대한 직접적인 증거는 아직 밝혀지지 않았습니다(플레밍 & 곤잘레스, 2009).
함께 고려하면 호르몬이 행동에 영향을 미치고 행동이 호르몬 분비에 영향을 미치기 위해 피드백을 주는 많은 예가 있습니다. 음식과 수분 섭취, 사회적 상호작용, 염분 균형, 학습과 기억, 스트레스 대처, 우울증, 불안 장애, 섭식 장애, 산후 우울증, 계절성 우울증을 포함한 정신병리 등 호르몬과 행동의 상호작용에 대한 예가 점점 더 많이 발견되고 있습니다. 추가 연구를 통해 이러한 호르몬과 행동의 상호작용이 어떻게 매개되는지 밝혀져야 합니다.
Outside Resources
- Book: Adkins-Regan, E. (2005). Hormones and animal social behavior. Princeton, NJ: Princeton University Press.
- Book: Beach, F. A. (1948). Hormones and behavior. New York: Paul Hoeber.
- Book: Beach, F. A. (1975). Behavioral endocrinology: An emerging discipline. American Scientist, 63: 178–187.
- Book: Nelson, R. J. (2011). An introduction to behavioral endocrinology (4th ed.). Sunderland, MA: Sinauer Associates.
- Book: Pfaff, D. W. (2009). Hormones, brain, and behavior (2nd ed.). New York: Academic Press.
- Book: Pfaff, D. W., Phillips, I. M., & Rubin, R. T. (2005). Principles of hormone/behavior relations. New York: Academic Press.
- Video: Endocrinology Video (Playlist) - This YouTube playlist contains many helpful videos on the biology of hormones, including reproduction and behavior. This would be a helpful resource for students struggling with hormone synthesis, reproduction, regulation of biological functions, and signaling pathways.
- https://www.youtube.com/playlist?list=PLqTetbgey0aemiTfD8QkMsSUq8hQzv-vA
- Video: Paul Zak: Trust, morality - and oxytocin- This Ted talk explores the roles of oxytocin in the body. Paul Zak discusses biological functions of oxytocin, like lactation, as well as potential behavioral functions, like empathy.
- Video: Sex Differentiation- This video discusses gonadal differentiation, including the role of androgens in the development of male features.
- Video: The Teenage Brain Explained- This is a great video explaining the roles of hormones during puberty.
- Web: Society for Behavioral Neuroendocrinology - This website contains resources on current news and research in the field of neuroendocrinology.
- http://sbn.org/home.aspx
Discussion Questions
- What are some of the problems associated with attempting to determine causation in a hormone–behavior interaction? What are the best ways to address these problems?
- Hormones cause changes in the rates of cellular processes or in cellular morphology. What are some ways that these hormonally induced cellular changes might theoretically produce profound changes in behavior?
- List and describe some behavioral sex differences that you have noticed between boys and girls. What causes girls and boys to choose different toys? Do you think that the sex differences you have noted arise from biological causes or are learned? How would you go about establishing your opinions as fact?
- Why is it inappropriate to refer to androgens as “male” hormones and estrogens as “female” hormones?
- Imagine that you discovered that the brains of architects were different from those of non-architects—specifically, that the “drawstraightem nuclei” of the right temporal lobe were enlarged in architects as compared with non-architects. Would you argue that architects were destined to be architects because of their brain organization or that experience as an architect changed their brains? How would you resolve this issue?
Vocabulary
- 5α-reductase
- An enzyme required to convert testosterone to 5α-dihydrotestosterone.
- Aggression
- A form of social interaction that includes threat, attack, and fighting.
- Aromatase
- An enzyme that converts androgens into estrogens.
- Chromosomal sex
- The sex of an individual as determined by the sex chromosomes (typically XX or XY) received at the time of fertilization.
- Defeminization
- The removal of the potential for female traits.
- Demasculinization
- The removal of the potential for male traits.
- Dihydrotestosterone (DHT)
- A primary androgen that is an androgenic steroid product of testosterone and binds strongly to androgen receptors.
- Endocrine gland
- A ductless gland from which hormones are released into the blood system in response to specific biological signals.
- Estrogen
- Any of the C18 class of steroid hormones, so named because of the estrus-generating properties in females. Biologically important estrogens include estradiol and estriol.
- Feminization
- The induction of female traits.
- Gonadal sex
- The sex of an individual as determined by the possession of either ovaries or testes. Females have ovaries, whereas males have testes.
- Hormone
- An organic chemical messenger released from endocrine cells that travels through the blood to interact with target cells at some distance to cause a biological response.
- Masculinization
- The induction of male traits.
- Maternal behavior
- Parental behavior performed by the mother or other female.
- Neurotransmitter
- A chemical messenger that travels between neurons to provide communication. Some neurotransmitters, such as norepinephrine, can leak into the blood system and act as hormones.
- Oxytocin
- A peptide hormone secreted by the pituitary gland to trigger lactation, as well as social bonding.
- Parental behavior
- Behaviors performed in relation to one’s offspring that contributes directly to the survival of those offspring
- Paternal behavior
- Parental behavior performed by the father or other male.
- Progesterone
- A primary progestin that is involved in pregnancy and mating behaviors.
- Progestin
- A class of C21 steroid hormones named for their progestational (pregnancy-supporting) effects. Progesterone is a common progestin.
- Prohormone
- A molecule that can act as a hormone itself or be converted into another hormone with different properties. For example, testosterone can serve as a hormone or as a prohormone for either dihydrotestosterone or estradiol.
- Prolactin
- A protein hormone that is highly conserved throughout the animal kingdom. It has many biological functions associated with reproduction and synergistic actions with steroid hormones.
- Receptor
- A chemical structure on the cell surface or inside of a cell that has an affinity for a specific chemical configuration of a hormone, neurotransmitter, or other compound.
- Sex determination
- The point at which an individual begins to develop as either a male or a female. In animals that have sex chromosomes, this occurs at fertilization. Females are XX and males are XY. All eggs bear X chromosomes, whereas sperm can either bear X or Y chromosomes. Thus, it is the males that determine the sex of the offspring.
- Sex differentiation
- The process by which individuals develop the characteristics associated with being male or female. Differential exposure to gonadal steroids during early development causes sexual differentiation of several structures including the brain.
- Target cell
- A cell that has receptors for a specific chemical messenger (hormone or neurotransmitter).
- Testosterone
- The primary androgen secreted by the testes of most vertebrate animals, including men.
References
- Alexander, G. M. & Hines, M. (2002). Sex differences in response to children’s toys in nonhuman primates (Cercopithecus aethiops sabaeus). Evolution and Human Behavior, 23, 467–479.
- Berenbaum, S. A., Martin, C. L., Hanish, L. D., Briggs, P. T., & Fabes, R. A. (2008). Sex differences in children’s play. In J. B. Becker, K. J. Berkley, N. Geary, E. Hampson, J. Herman, & E. Young (Eds.), Sex differences in the brain: From genes to behavior. New York: Oxford University Press.
- Dabbs, J. M. (2000). Heroes, rogues, and lovers: Testosterone and behavior. Columbus, OH: McGraw Hill.
- Fleming, A. S., & Gonzalez, A. (2009). Neurobiology of human maternal care. In P. T. Ellison & P. B. Gray (Eds.), Endocrinology of social relationships (pp. 294–318). Cambridge, MA: Harvard University Press.
- Fleming, A. S., Steiner, M., & Corter, C. (1997). Cortisol, hedonics, and maternal responsiveness in human mothers. Hormones and Behavior, 32, 85–98.
- Goodson, J. L., Saldanha, C. J., Hahn, T. P., Soma, K. K. (2005). Recent advances in behavioral neuroendocrinology: Insights from studies on birds. Hormones and Behavior, 48, 461–73.
- Kidd, K. A., Blanchfield, P. J., Mills, K. H., Palace, V. P., Evans, R. E. Lazorchak, J. M. & Flick, R. (2007). Collapse of a fish population following exposure to a synthetic estrogen. Proceedings of the National Academy of Sciences,104, 8897–8901.
- Nelson, R. J. (Ed.) (2006). Biology of aggression. New York: Oxford University Press.
- Nelson, R.J. (2011). An introduction to behavioral endocrinology (4th ed.). Sunderland, MA: Sinauer Associates.
- Phoenix, C. H., Goy, R. W., Gerall, A. A., & Young, W. C. (1959). Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology, 65: 369–382.
- van Anders, S., Hamilton, L., Schmidt, N., & Watson, N. (2007). Associations between testosterone secretion and sexual activity in women. Hormones and Behavior, 51, 477–82.
Authors
 Randy J. NelsonRandy J. Nelson has two PhDs (Psychology and Endocrinology) and is Professor and Chair of Neuroscience and Professor of Psychology at The Ohio State University. He has been appointed as University Distinguished Professor for his work on hormones, biological rhythms, and behavior. Dr. Nelson has published nearly 400 papers and several books on these topics.
Randy J. NelsonRandy J. Nelson has two PhDs (Psychology and Endocrinology) and is Professor and Chair of Neuroscience and Professor of Psychology at The Ohio State University. He has been appointed as University Distinguished Professor for his work on hormones, biological rhythms, and behavior. Dr. Nelson has published nearly 400 papers and several books on these topics.
Creative Commons License



 Hormones & Behavior by Randy J. Nelson is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Permissions beyond the scope of this license may be available in our Licensing Agreement.
Hormones & Behavior by Randy J. Nelson is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. Permissions beyond the scope of this license may be available in our Licensing Agreement.